2023.08.11
【プレスリリース】ソバゲノムの解読 ―高精度ゲノム解読がソバの過去と未来を紡ぐ―
概要
2050年の世界人口は97億と予想され、イネ、コムギ、トウモロコシなどの三大穀物への食料の依存が問題視されています。これに対し、食料としての価値が高いにもかかわらず研究が遅れ、未開発のポテンシャルが秘められたままの「孤児作物」への関心が増しています。次世代シーケンシング 1) 技術による孤児作物のゲノム解読は、その効率的な育種を促進し、飢餓の撲滅や栄養改善などのSDGs達成への重要なステップとなることが期待されています。
安井康夫(京都大学大学院 農学研究科・助教)らの国際共同研究グループ(理研・農研機構・千葉大学・京都府立大学・かずさDNA研究所・総合研究大学院大学・雲南農業大学・ケンブリッジ大学など)は、孤児作物の一つであるソバのゲノム配列を染色体レベルで高精度に解読することにより、ソバのゲノムの進化と栽培ソバの起原を解き明かしました。さらに、予測された遺伝子をゲノム編集技術に依存しない手法で改変しました。その結果、これまで世界に存在しなかったモチ性ソバを開発することに成功しました。またソバの繁殖様式を他殖性から自殖性へ転換させることにより、新たな自殖性ソバの開発にも成功しました。本研究で用いられた育種方法は、ゲノム編集技術に未対応な多種多様な孤児作物の改良に貢献することが期待されます。
本成果は、現地時間2023年8月10日午後4時に英国の国際学術誌「Nature Plants」にオンライン掲載されます。

1. 背景
2050年には、世界人口は97億人に達すると予想されています。現在、世界の人間のカロリー需要の多くは、いわゆる三大穀物とよばれるイネ(Oryza sativa)、コムギ(Triticum aestivum)、そしてトウモロコシ(Zea mays)によって供給されています。今後の地球温暖化の進行がこれら三大穀物の収穫量の増加率を低下させる懸念があり、食料安全保障を三大穀物にのみ依存する考え方が問われています。このため今後、農業生産システムには大きな変革が訪れると予見されています。
ソバなどの地域的または伝統的な重要性を持つ多くの作物では、商業価値が低いためにその育種学的研究が立ち遅れていました。このような作物は「孤児作物」と呼ばれています。孤児作物の育種を推し進め、その作物としてのポテンシャルを引き出すことは、世界的に食品の多様化を促進し、栄養不足を解消します。また、各地域でよく根付いている孤児作物の収量増大は、農家の収入向上をもたらすとともに、食料問題の解決にも貢献します。すなわち、孤児作物の改良は、国連の持続可能な開発目標(SDGs)の中で、特に目標2(飢餓を終わらせ、食料安全保障及び栄養改善を実現し、持続可能な農業を促進する)の実現に向けた重要な研究と位置付けられています。近年の次世代シーケンシング技術の進歩とその汎用化は、孤児作物においてもゲノム解読を可能としており、これまでの孤児作物での育種学的研究の遅れを取り戻すことができると期待されています。実際に近年では、キヌアやフォニオ、シコクビエ、ホースグラムなどの多くの孤児作物のゲノム解読が行われています。ゲノム科学を活用することで孤児作物の改良を高効率で達成することが世界における育種学の大きな流れになっています。
ソバ(学名: Fagopyrum esculentum ssp. esculentum)の子実はエネルギー源となるデンプンの他、ビタミン、ミネラル、食物繊維を多く含んでおり、栄養価の高い食品として知られています。ソバは日本においては主に麺(蕎麦切り)として食されていますが、アジアの多くの国々でも麺やパンとして食されています。またアジア以外でもフランスのガレットやイタリアのピッツォッケリなど、ヨーロッパにおいてもソバは伝統的に利用されており、世界の温帯地域を中心に広く栽培されています。このようにソバは世界で広く利用されているものの、私たちは日本の食文化を象徴するソバ研究の道筋を切り開く役割は、なんとしても日本の研究者が担うべきであると考え、2016年に世界に先駆けてソバのドラフトゲノム配列 2) (概要ゲノム配列)を公開しました。しかしながら、ドラフトゲノム配列は断片的であり、連続性に欠けているため、正確な遺伝子の予測が困難であり、また各遺伝子がどの染色体にどの程度の距離で位置しているか、という重要な情報が欠けていました。
今回、私たちは、この問題を解決するため染色体レベルでの高精度なソバのゲノム配列の解読に取り組みました。ゲノム配列は生命の設計図であり、そこには生命の歴史が刻まれています。すなわちゲノムを解読することにより、生命の「過去」の歴史を紐解くことができます。また同時に設計図を用いて、生物や作物を改良し、人類の「未来」を拓くことができます。
そこで私たちはソバの高精度ゲノム配列を利用して、ソバのゲノムの進化と栽培ソバの起原の解明を目指しました。さらに高精度ゲノム配列から予測された遺伝子をゲノム編集技術に依存しない汎用性の高い手法で改変し、これまで世界になかったモチ性ソバの開発や新たな自殖性ソバの開発に挑みました。モチ性ソバとは、もちもちとした食感を持つ新たなソバの形質です。イネ、ムギ、トウモロコシ、アワ、ヒエなどの多くの穀類にはモチ性形質が存在しますが、ソバにはこれまで存在しなかったものです。日本の伝統的な食文化として、年越しには蕎麦を食べ、新年には餅を食べるという風習がありますが、モチ性のソバがなかったのは不思議なことです。ソバとモチ、この2つを組み合わせることで、新たな食文化の発展が期待できると考えられます。具体的には、モチ性がある蕎麦は、麺の歯応えを向上させ、さらに製麺時につなぎの使用を減らす可能性があります。これにより、十割蕎麦の製造がより容易になると期待できます。また、京都をはじめとした地域ではソバを使った多くの和菓子がありますが、モチ性により新たな和菓子を創出することも可能です。さらに、現在ではソバは主に粉にして食べられていますが、モチ性のあるソバの実には弾力と粘り気があり、粒食としてお箸で食べることも可能になります。このモチ性ソバの開発は、ソバのような孤児作物においても、今までになかった形質を持つ品種育成を可能とする、すなわち作物の秘められたポテンシャルを解放する好例となります。そこで新たな食文化の創造だけでなく、多様な孤児作物の形質改良に対する新たな道を切り開く可能性を世界に示すため、その象徴的な成果として世界初となるモチ性ソバの開発に挑みました。また、モチ性ソバの開発に利用した変異導入技術は、学術的な観点からも有用であり、この一環としてソバの自家不和合性 3) を制御する遺伝的機構の解明にも取り組み、ソバの生殖様式を他殖性から自殖性へと転換させて、新たな自殖性ソバの開発にも挑みました。
2. 研究手法・成果
【ソバゲノム参照配列 4) の作成】
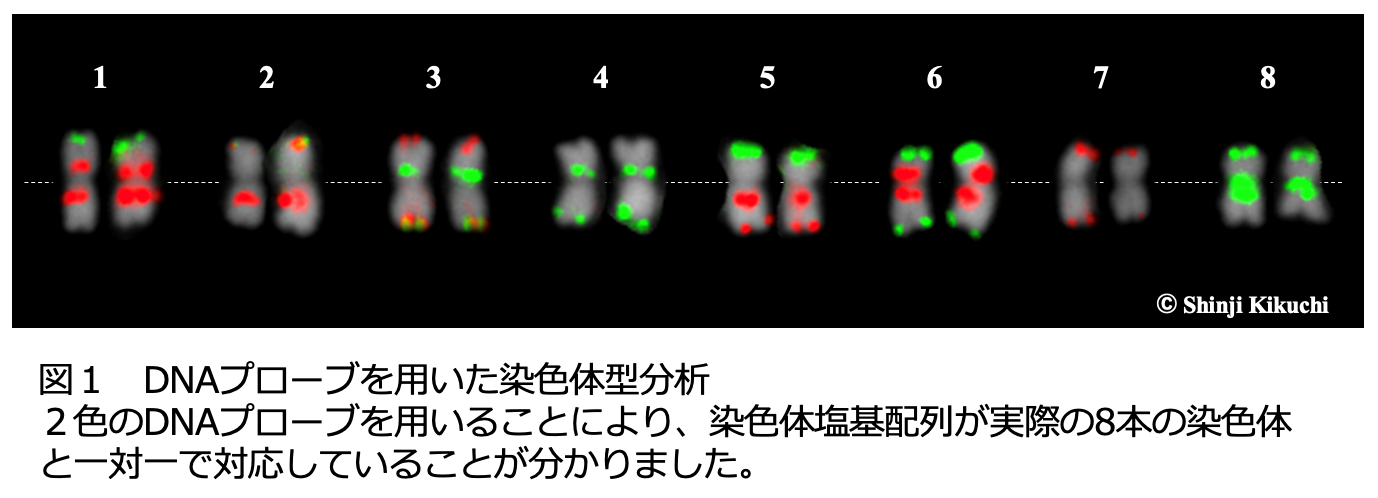
ソバは16本の染色体(2n = 2x =16)を持ち、そのゲノムサイズは約1.27 Gb(12億7千万塩基)です。私たちはまず、最新のゲノム解読技術を用いてソバの全染色体の塩基配列を解読しました。この染色体塩基配列の一部を蛍光標識したDNA(プローブDNA)を合成し、FISH法 5) を用いて染色体塩基配列が実際の染色体と一対一に対応することを顕微鏡で確認しました(図1)。この配列はソバゲノム全体の約96%をカバーしており、今後のソバの遺伝学的・育種学的研究のための参照配列と位置づけました。また参照配列を解析した結果、合計30,608個の遺伝子を確認できました。
【全ゲノム重複 6) の発見】
ゲノムにはその生物の「過去」の進化の歴史が刻まれており、ゲノムデータを用いた数理科学・情報科学解析によってそれを読み解くことができます。私たちはまず、ソバゲノム中に配列の似た遺伝子対がたくさん見られることに着目し、他の植物種10種の全遺伝子データを加えた比較ゲノム・分子進化解析を行いました。その結果、ソバを含むタデ科植物の共通祖先において、約7,100万年前と8,500万年前の2回にわたり、全ゲノム重複という現象が起きていたことが示されました。全ゲノム重複ではゲノム全体が倍になるため、全ての遺伝子につき、配列が同じ遺伝子が新しく作られます。この2回の全ゲノム重複の結果、約7,100万年たった今でも、たくさんの配列の似た重複遺伝子対がゲノム中に残っていたのです。ソバ以外にも多くの植物が、恐竜を含む生物の大量絶滅が生じた約6,000-7,000万年前に全ゲノム重複を経験しており、この時代に生じた全ゲノム重複が現在の植物の多様性に大きく貢献したと考えられています。
【栽培ソバの起原】
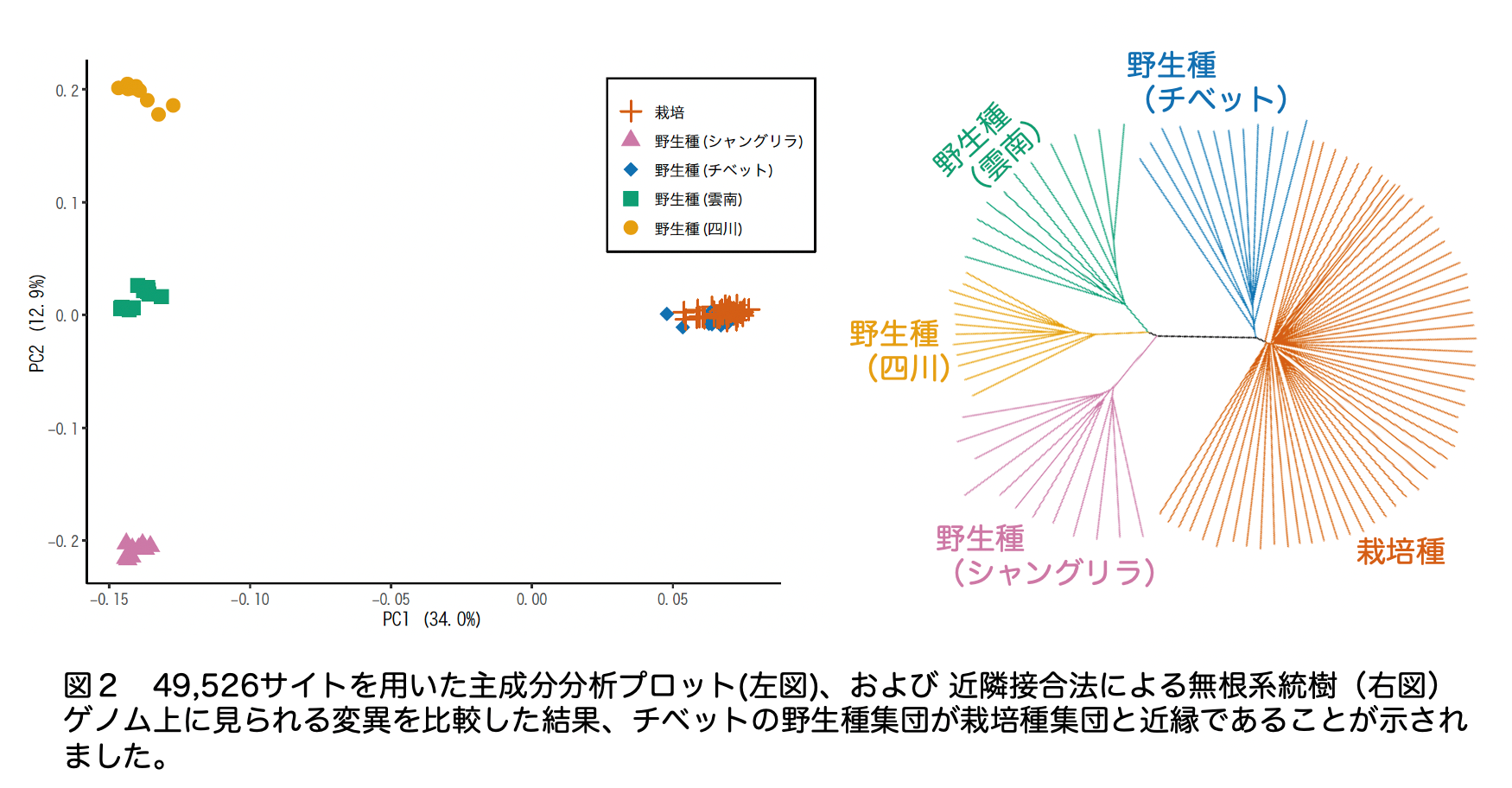
私たちが食している栽培作物は「過去」に自然界の野生種から人類が選抜し、栽培を始めたことで起原しました。その起原地と祖先となる野生種の探索は、人類の歴史の理解や、育種の際の有用遺伝子の同定につながります。これまでに栽培ソバの祖先野生種(F. esculentum ssp. ancestrale)が中国の雲南省、四川省、およびチベット自治区の境界周辺に自生していること、そして、チベット南東部の祖先野生種集団が栽培種と近縁であることが、京都大学名誉教授の大西近江博士らにより明らかにされていました。本論文の著者の多くは大西博士の研究室の出身であり、私たちは大西博士の結果をゲノムレベルで確認し、さらにゲノムに刻まれた栽培化時の人為選択の検出に挑みました。そして栽培ソバ(57個体)と祖先野生種(47個体)のゲノムのショートリード 7) を解読し、これらを参照配列と比較することにより、チベット南東部の祖先野生種の集団が栽培種と最も近縁であることを確認しました(図2)。これにより先端的な情報技術を活用し、大西博士の説が裏付けられたことになります。さらに、栽培種の第一染色体には明瞭な人為選抜の痕跡がみられ、チベット南東部の祖先野生種の集団から選抜された特定の遺伝子型が世界中の栽培ソバに広がっていることが明らかになりました。これは、ただ単に栽培種とチベット南東部の祖先野生種集団のゲノム全体の塩基配列が似ているだけではなく、人為選抜の対象となった遺伝子型を持つ集団がチベット南東部に存在したことを示しています。このため私たちは、栽培ソバの起原としてチベット南東部の集団が関与したと結論づけました。
【世界初となるモチ性ソバの育成】
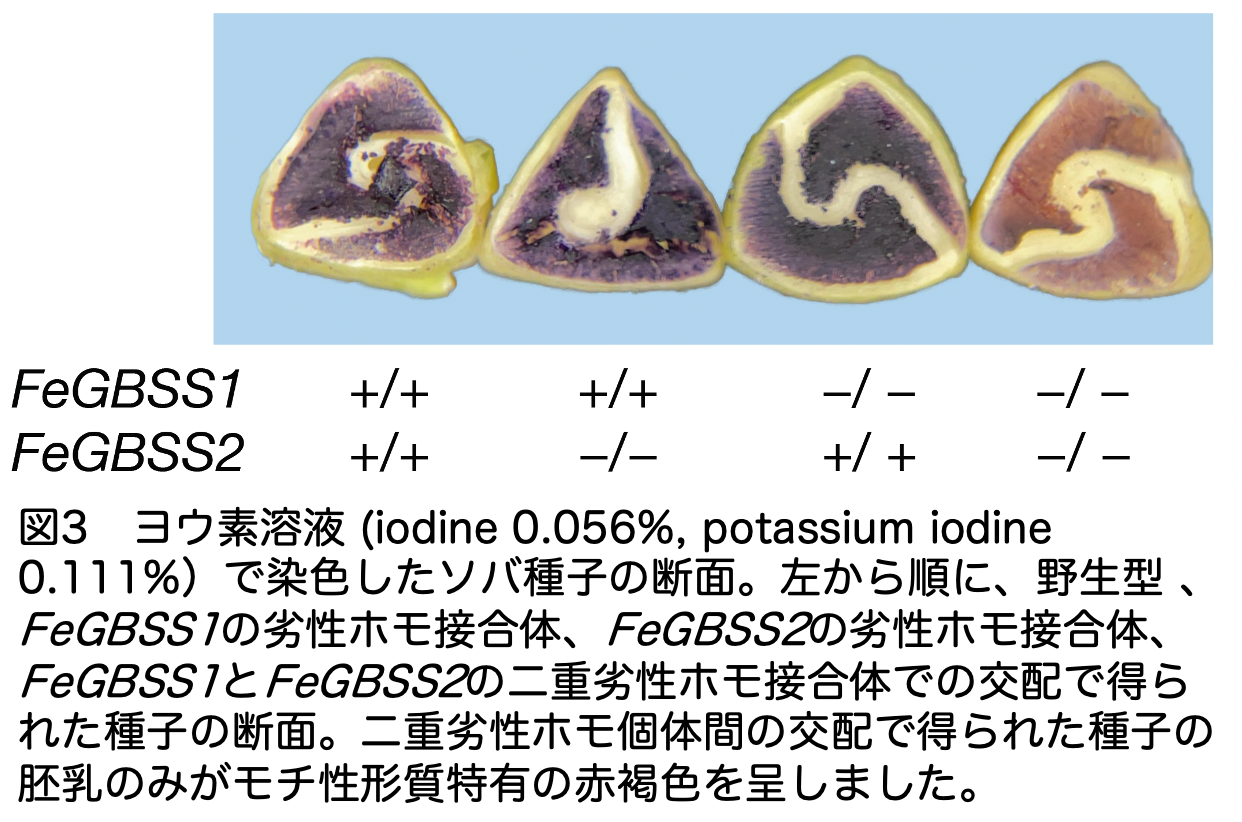
日本の年末年始には、年越し蕎麦を食べ、翌朝には新年の挨拶と共に餅を食べます。蕎麦と餅、これらは両方とも日本人にとって重要な食品でありながら、何故かこれまでのソバ栽培の歴史においてモチ性のソバは見出されていませんでした。イネなどでは、顆粒結合型デンプン合成遺伝子(Granule bound starch synthase; GBSS)が機能しなくなるとモチ性が顕れることが既知です。そして、多くの穀類ではこれまでの栽培化の歴史の中でモチ性、すなわち機能を失ったGBSSが選抜されてきました。そこで私たちは、ソバが2度の全ゲノム重複を経験した事実を考慮し、1)ソバのゲノムに複数のGBSSが存在し、2)これら全ての遺伝子の機能が同時に失わなければモチ性は顕れないと予測しました。すなわち、全ての遺伝子が同時に機能を失う確率は非常に低いと考えたのです。参照配列を解析した結果、実際にソバのゲノムには5つのGBSSが存在していました。更に、これら5つのうち2つ(FeGBSS1とFeGBSS2)がソバの種子胚乳、つまり食用となる部分において高いレベルで発現されていることがわかりました。これらの発見から、FeGBSS1とFeGBSS2の機能を抑制すれば、モチ性ソバを作り出せると考えました。しかし、ソバは他の多くの孤児作物と同様、ゲノム編集技術の開発がまだ十分に進んでいません。この問題に対応するため、私たちは、様々な作物の品種改良に利用されてきた化学物質、エチルメタンスルホン酸(EMS)を用いることにより、5,801個体からなるソバの変異誘導集団を作り出しました。そして、次世代シークエンサーを使った大規模な変異検出解析から、機能が損なわれたFeGBSS1とFeGBSS2を見出しました。その後、ヨウ素デンプン反応を用いてこれら変異遺伝子を持つ植物体の胚乳を調査したところ、FeGBSS1あるいはFeGBSS2のどちらか一方だけの機能を失った場合にはモチ性は顕れず(青色を呈する)、それらの遺伝子機能が両方同時に喪失したときに初めてモチ性が顕れることが明らかになりました(赤紫色を呈する)(図3参照)。これらの結果から、モチ性ソバがこれまで発見されなかった主な理由は、7,100から8,500万年前の全ゲノム重複によるGBSS遺伝子のコピー数の増加であると考えられました。このようにモチ性ソバの開発では、ゲノム解析によって「過去」の遺伝子重複の謎が解け、それが新たな「未来」、つまりこれまでに存在しなかったモチ性ソバの開発へとつながりました。
【ソバの自家不和合性遺伝子の同定】
ソバには「雌しべが短く、雄しべの長い花型(短柱花)」を持つ個体と「雌しべが長く、雄しべの短い花型(長柱花)」を持つ個体が存在し、異なる花の形を持つ個体間でしか交配できません。このような生殖様式は二花柱性と呼ばれ、ソバ以外にも200種以上の植物で見られます。二花柱性は、雄しべ・雌しべの長さと自家不和合性を制御する複数の遺伝子が隣接し、1つの遺伝子のように振る舞うことで成立しています。このような遺伝子群は「スーパージーン」と呼ばれ、哺乳類・鳥類の性決定やチョウの擬態など、多くの複雑な形態に関与しています。これまでにすでに私たちは、二花柱性に関連する遺伝子として、S-LOCUS EARLY FLOWERING 3(S-ELF3)を同定しており、今回の高精度ゲノム配列の結果からもS-ELF3が二花柱性を制御するスーパージーンの構成因子であることが確認できました。さらに私たちは、S-ELF3が雄しべや雌しべの長さなどの花の形態を制御しているのか、それとも自家不和合性を制御しているのかを明らかにするため、EMSを用いてS-ELF3の機能を欠損させました。その結果、欠損型のS-ELF3を持つ個体では雌しべの長さが長くなり、さらに自殖することが分かりました(図4)。これはS-ELF3が短柱花の雌しべの長さと自家不和合性の形質を同時に制御していることを示しています。それぞれ独立に二花柱性が進化したPrimula属やTurnera属植物でも同様に、1つの遺伝子が短柱花の雌しべの長さと自家不和合性の形質を同時に制御していることから、二花柱性の進化における共通の原理原則がうかがえます。今回の結果は、生物の複雑な生殖様式やスーパージーンの進化を理解する上での重要な一歩だと考えています。
3.波及効果、今後の予定
4.研究プロジェクトについて
MEXT | Japan Society for the Promotion of Science (JSPS) - 20K06761 [フォーセット]
MEXT | Japan Society for the Promotion of Science (JSPS) - 21H00356 [フォーセット]
MEXT | Japan Science and Technology Agency (JST) - JPMJAX20BA [竹島]
Leverhulme Trust - RPG-2017-196 [ジョーンズ]
NARO | Bio-oriented Technology Research Advancement Institution (BRAIN) - JPJ007097 [原]
MEXT | Japan Society for the Promotion of Science (JSPS) - 22H05181 [白澤]
MEXT | Japan Society for the Promotion of Science (JSPS) - 18KK0172 [安井]
Cabinet Office, Government of Japan - JPJ009237 [安井]
用語解説
- 1) 次世代シーケンシング(Next-Generation Sequencing; NGS)
DNAの塩基配列を並列に、つまり一度に大量の塩基配列を高速に解読する技術です。これにより、生物のゲノム解読が可能となり、また遺伝子の変異や構造変化など、DNAレベルでの詳細な情報を短時間で解析することも可能となります。 - 2) ドラフト配列
生物のゲノムの初期的な配列情報のことを指します。これは、生物のゲノム配列を初めて解読する際の一種の草案で、短いゲノム配列断片の集合体として存在します。しかし、これらの断片がゲノム全体の中でどのような配置になっているのかは明らかではないため、遺伝子の正確な予測やそのゲノム上での正確な位置を特定することが困難です。 - 3) 自家不和合性
受粉後に花粉(もしくは花粉管)と雌しべの間で自己認識が生じ、自己の花粉管の伸長が抑制される現象。このため、自家不和合性植物は自家受精ができず、他殖性となります。裸子植物から被子植物へと進化した後に獲得された性質であり、被子植物の半数以上がこのような性質を持つとされています。 - 4) 参照配列
ゲノムの参照配列とは、特定の生物種のゲノムの詳細なゲノム配列情報のことを指します。これは通常、他の個体のゲノム配列を解析する際の基準として用いられます。これにより、新しく得られたゲノム配列の特徴や変異を比較・解析することが可能となります。 - 5) FISH
蛍光 in situ ハイブリダイゼーション(Fluorescence In Situ Hybridization)。染色体の標的DNA配列とそれに相補的なプローブDNAを結合(ハイブリダイズ)させ、そのプローブDNAを蛍光検出することで染色体上の標的DNA配列の位置や数を可視化する方法です。 - 6) 全ゲノム重複
ゲノム全体が倍加する現象で、植物だけでなく、動物や酵母などの生物でも観察されています。特に植物では全ゲノム重複が頻繁に生じており、新しい種の誕生や既存種の適応能力向上に寄与し、進化の過程で重要な役割を果たしてきたと考えられています。 - 7) ショートリード
次世代シーケンシング(NGS: Next Generation Sequencing)技術によって得られる、比較的短い長さのDNA配列。本研究では300 bpの長さのDNA配列を大量に並列に読み取り、参照配列との比較に用いました。
研究者のコメント
今回の研究成果は農学、食品科学、そしてゲノム研究の境界を押し広げるものだと思っています。ここから新たな食文化が生み出され、ソバの利用範囲が拡大されると期待しており、とてもワクワクしています。また、同様の研究が多様な孤児作物に展開され、新たな段階への一歩となることを熱望しています。私たちの目標は「孤児作物の可能性を解き放ち、世界の食料問題に挑む!」です。
論文タイトルと著者
参考図表